SET1ドメインタンパク質SUVH2によるトランスポゾンの制御機構を解明 ~環境ストレス応答で活性化するトランスポゾンの制御機構についての新しい知見~
SET1ドメインタンパク質SUVH2によるトランスポゾンの制御機構を解明 ~環境ストレス応答で活性化するトランスポゾンの制御機構についての新しい知見~
ポイント
・SUVH2タンパク質を欠失させると、ONSENの転写量が上昇。
・SUVH2タンパク質を欠失させても、ONSENの転移は見られない。
・siRNAがONSENの転移制御に重要な役割。
概要
北海道大学大学院理学研究院の伊藤秀臣准教授の研究グループは、環境ストレスで活性化するシロイヌナズナの転移因子(トランスポゾン)の制御機構について明らかにしました。 研究グループは、環境ストレス、特に熱ストレスで活性化するトランスポゾン(ONSEN)に着目し、その活性を制御する植物側の因子について長年研究してきました。その中で、ONSENはRNA誘導型のDNAメチル化 (RdDM*1)によって転写活性が制御されていることを明らかにし、RdDMがうまく機能しないような変異体では、ONSENの転写活性が増加し、次の世代にONSENが転移することも分かりました。RdDM経路の因子の一つであるSUPPRESSOR OF VARIEGATION 3–9 HOMOLOG (SUVH)ファミリータンパク質の一つSUVH2の変異体に高温ストレスを与えると、他のRdDM経路の変異体同様、ONSENの高発現が見られました。 ところが、興味深いことに、SUVH2の変異体ではONSENの世代を超えた転移は観察されませんでした。本研究では、SUVH2変異体ではONSENの配列相補的な小分子RNA(siRNA*2)が生成されることが分かりました。 一方、ONSENの世代を超えた転移が観察される変異体ではsiRNAが検出されませんでした。このことから、siRNAがONSENの転移を制御していることが示唆されます。今回の研究結果から、高温で活性化するトランスポゾンONSENの制御には転写レベルでの制御と転移レベルでの制御が独立に存在していることが明らかになりました。そして、転移の制御にはsiRNAが重要な役割を担っているという新しい知見を得ることができました。 なお、本研究成果は、2024年2月8日(水)公開のFrontiers in Plant Science誌に掲載されました。また、北大プレスリリース<https://www.hokudai.ac.jp/news/2024/02/set1suvh2.html>にも掲載されております。
【背景】
転移因子(トランスポゾン)はあらゆる生物に存在する“動く遺伝子”であり、トランスポゾンの転移はゲノムに悪影響を与える可能性があります。そのため、トランスポゾンの転移制御は、宿主にとって自身のゲノムの安定性を守るために重要です。ほとんどのトランスポゾンはDNAのメチル化やヒストン修飾などのエピジェネティックな修飾により転写が抑制されているため、その転移制御機構の詳細は明らかになっていませんでした。
シロイヌナズナにおける熱活性型レトロトランスポゾンONSENは37℃の熱ストレスで転写が活性化し、染色体外DNAを産生します。siRNAを介したトランスポゾンの転写制御機構であるRdDM経路の変異体では、熱ストレスによりONSENの世代を超えた転移が観察されました。
これまでの報告から、シロイヌナズナには10種類のsuppressors of variegation 3-9 homolog (SUVH)ファミリータンパク質が存在し、そのうちSUVH2とSUVH9がRdDMのDNAメチル化活性と密接に関連していることが示されていました。この研究では、レトロトランスポゾンのサイレンシングのメカニズムにおけるSUVH2の役割を解析しました。
【研究成果】
本研究では、SUVH2変異体におけるONSENの活性制御について解析しました。SUVH2の機能を欠損させると、ONSENの転写量が上昇しました。
次に、世代を超えたONSENの転移活性を調べました。RdDM経路の変異体では、世代を超えたONSENの転移が観察されますが、SUVH2変異体ではONSENの転移は観察されませんでした。転移が観察されなかった原因を調べるために、ONSENのDNAメチル化を調べました。その結果、ONSEN配列上のDNAメチル化は野生型と比較して低下していました。このDNAの脱メチル化は、ONSENの転写活性と相関があることが分かりました。
さらに、クロマチンの凝縮度合いがONSENの転移と関係があるかどうか調べるために、SUVH2変異体におけるオープンクロマチン状態をFAIRE-qPCR法とDAPI染色法を用いて解析しました。その結果、SUVH2変異体は野生型と同様のクロマチン状態であることが分かり、ONSENの転移とクロマチンの凝縮の関連性は見られませんでした。次に、SUVH2変異体における小分子RNAの蓄積量を調べたところ、ONSEN配列と相補的な小分子RNAの蓄積が検出されました。
一方、ONSENの世代を超えた転移が見られるRdDM経路の変異体ではこの小分子RNAが見られないことから、ONSEN由来の小分子RNAが世代を超えたONSENの転移制御に関与していることが示唆されました。
以上の結果から、SUVH2は熱ストレスで活性化するONSENの転写活性制御に重要であるが、転移制御には小分子RNAを介した機構が関与していることが明らかになりました(図1)。
【今後への期待】
この研究では環境ストレスで活性化するトランスポゾンと宿主の関係について、エピジェネティックな因子が重要な役割を果たしていることを明らかにしました。トランスポゾンの制御には転写の制御と転移の制御が存在します。転写制御についてはDNAのメチル化やヒストン修飾が重要であることが分かってきましたが、転移制御については、まだ分からないことだらけです。
今回明らかになった小分子RNAがどのようにトランスポゾンの転移を制御しているのか今後さらに研究が必要です。
【謝辞】
本研究は、日本学術振興会科学研究費助成事業・国際共同研究加速基金(国際共同研究強化(A)(研究代表:伊藤秀臣、課題番号21KK0263)、日本学術振興会科学研究費助成事業・基盤研究(C)(研究代表:伊藤秀臣、課題番号21K06008)、アンビシャス博士人材フェローシップ(研究代表:牛 小蛍)の助成を受けた成果です。
論文情報
論文名 Regulatory mechanism of heat-active retrotransposons by the SET Domain Protein SUVH2(SETドメインタンパク質SUVH2による熱活性レトロトランスポゾンの制御機構)
著者名 牛 小蛍1,葛 智宇1,伊藤秀臣2*(1北海道大学大学院生命科学院,2北海道大学大学院理学研究院 *責任著者)
雑誌名 Frontiers in Plant Science(オープンアクセス科学ジャーナル)
DOI 10.3389/fpls.2024.1355626
公表日 2024年2月8日(水)(オンライン公開)
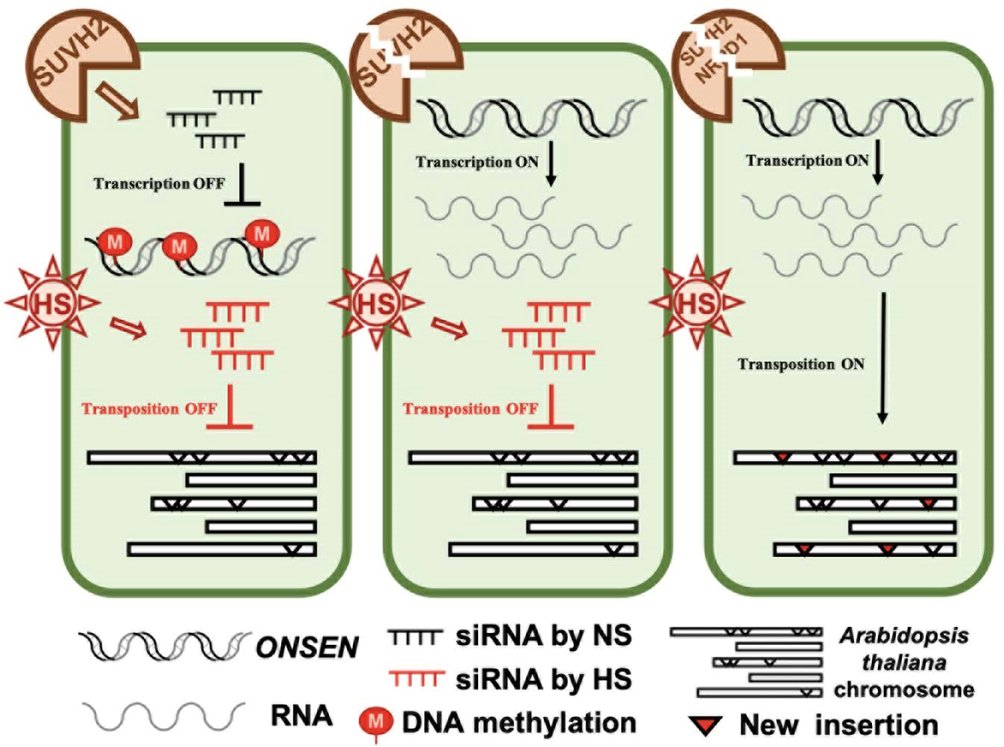
図1. ONSENのサイレンシングのモデル。
野生型では、RdDM経路でsiRNAが産生され、ONSENの転写を抑制する。suvh2変異体では、RdDM経路が破壊され、熱ストレスによってONSENの転写が解放される。熱ストレス下では、植物はSUVH2に依存する経路をバイパスして新しいsiRNAを産生し、熱ストレスによって産生されたこれらのsiRNAは、ONSENの転移を抑制する。suvh2/nrpd1二重変異体では、熱ストレス条件下でsiRNAを合成することができず、ONSENの転移制御が解除される。
【用語解説】
*1 RdDM … RNA指令型DNAメチル化機構。植物で最初にみつかった機構で小分子二本鎖RNAを介してDNAのメチル化が誘導される。メチル化DNAはヒストン修飾と共にテロクロマチン化を促進し、トランスポゾンや反復配列の制御に重要な役割を担っている。なお、DNAのメチル化とは、DNAメチル化酵素の働きにより、DNAを構成するA、T、G、Cの塩基のうち主にC(シトシン)のピリミジン環の5位炭素原子にメチル基修飾が付加される化学反応のことを言う。
*2 siRNA … 21-24塩基対から成る低分子二本鎖RNA。 siRNAはRNA干渉(RNAi)と呼ばれる現象に関与しており、伝令RNA(mRNA)の破壊によって配列特異的に遺伝子の発現を抑制する。植物においてはRdDM経路を介してDNAの新規メチル化を誘導する。