昆虫脳内における機械感覚刺激方位情報の階層的処理機構を明らかに
大学院生命科学院生命システム科学コース博士後期課程3年の千田輝さんと行動神経生物学系の小川宏人教授は、北里大学データサイエンス学部の設樂久志講師とともに、コオロギ脳内における気流応答性ニューロンの網羅的な電気生理学的解析を行い、気流機械感覚の神経情報処理機構を明らかにしました。本研究成果は、Journal of Neurophysiology誌に2025年9月18日オンライン公開、10月16日に掲載されました。
動物にとって刺激がどちらからやってくるかという方位情報は、その刺激を発する刺激源、例えばエサ、繁殖相手、あるいは捕食者がどちらに位置しているかを認識する上で,非常に重要な手がかりとなります。そしてその情報をもとに、動物は刺激源に近づく、あるいは遠ざかるといった定位行動を示します。このような目標指向型行動(Goal-directed behavior)では、刺激方位の認識とそれに基づいた移動運動の企画、そして運動の制御という3つの課題が脳によって解かれなければなりません。目標指向型行動はあらゆる動物で見られる本質的な行動ですが、実は刺激方位の認識〜運動企画〜運動制御のすべての過程をつなぐ神経回路と演算内容が明らかにされたものはほとんどなく、それも刺激に対して常に同じ定型的な運動を繰り返すような、ごく単純な反射的な行動にすぎません。そこで小川教授らのグループではコオロギの気流誘導性逃避行動をモデルとして、その課題に取り組んでいます。
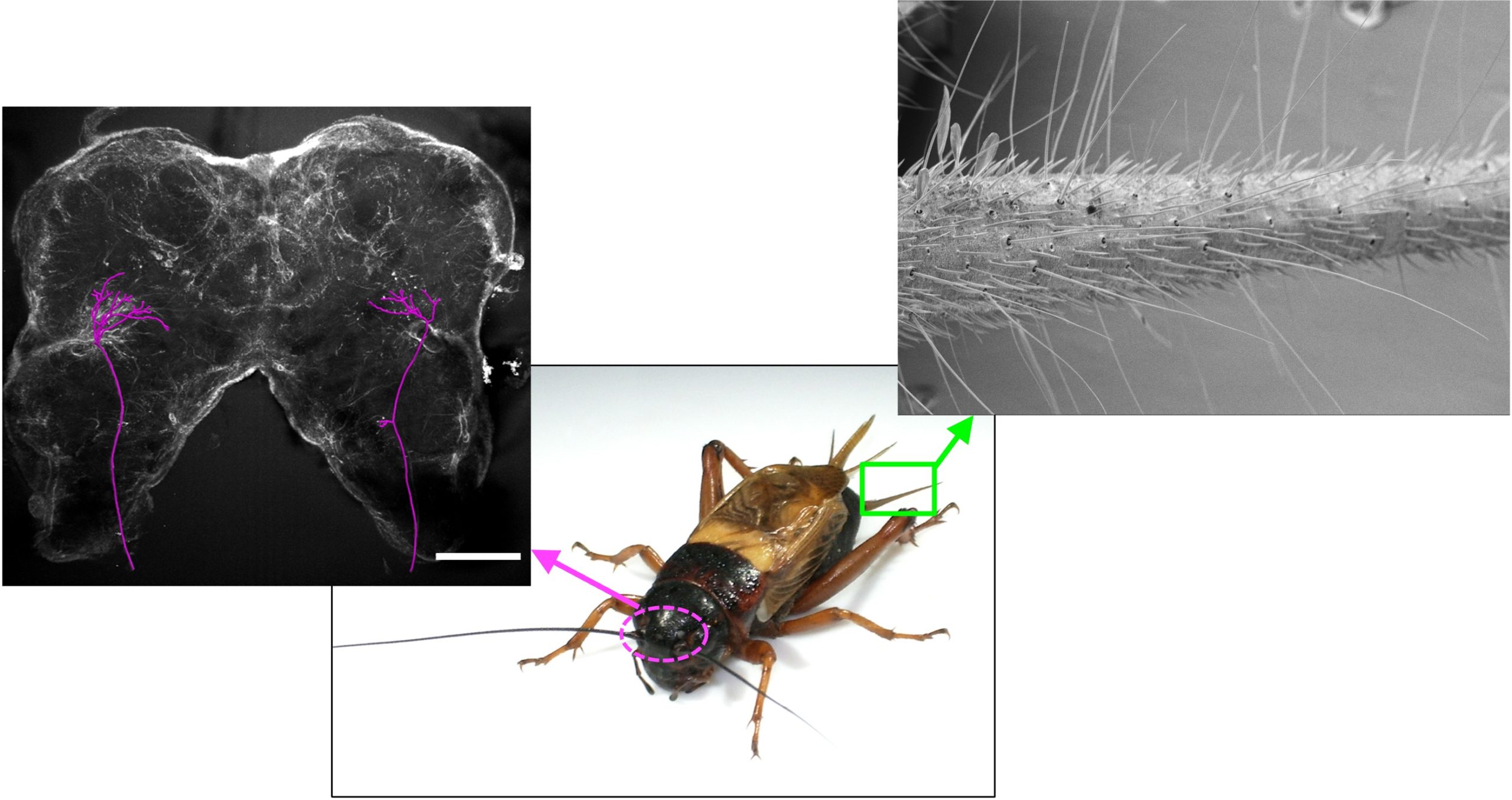
図1 フタホシコオロギ(中央)の脳(右)と尾葉(右)。脳内のマゼンダのトレースは気流情報を脳に運ぶ巨大介在ニューロンの神経軸索投射。
コオロギは短い気流刺激を受けると、刺激の到来と反対方向に走ったり、ジャンプしたりして遠ざかろうとする運動を示します。これは気流刺激を捕食者の接近を示唆する脅威信号と捉え、それから逃れようとする防御行動の一つであると考えられています。古典的な神経行動学の枠組みでは、このような逃避行動は上述したような生得的な反射行動であり、ごく少数のニューロンから構成される単純な神経回路によって実行されていると考えられてきました。しかし近年、逃避行動は単純な反射行動ではなく、捕食者までの距離や方向、周囲の環境、さらに被食動物の内的な状態によって柔軟に変化する行動であり、神経科学において様々な研究課題に取り組む良いモデルであると再認識されるようになりました。小川研究室でも、これまで様々な行動実験によって、コオロギの気流誘導性逃避行動が気流刺激の様相や周囲の環境、運動状態によって変化するものであることを報告しています。また、逃避行動の実行には脳からの下行性神経信号が必要であることもわかりました。
これまで小川教授らのグループは目標指向型行動の最初のステップ、すなわち気流刺激の方位をどのように検出し、脳に伝えているか、という課題について神経生理学的な研究を行ってきました。気流刺激はコオロギの腹部末端にある一対の尾葉と呼ばれるアンテナ状の機械感覚器官で受容されます(図1右)。尾葉上には片側あたり500〜700本の毛状機械感覚子という受容器があり、その毛状の突起に空気分子が衝突することで気流を検出します。毛状突起は根元のソケットにある蝶番状構造によって特定の方向に倒れやすくなっており、その蝶番状構造の向きによって、受容器が最も反応する気流の方向が決定されます。言い換えると毛によってそれぞれ気流の方向に対する感受性が異なるのです。毛状機械感覚子の感覚ニューロンからの信号は最終腹部神経節に伝えられ、その内部の神経回路によって気流の強度や方向、周波数などの情報が抽出されます。抽出された気流情報は、巨大介在ニューロン(Giant Interneurons: GIs)と呼ばれる上行性投射ニューロンによって脳や胸部神経節などの上位中枢へと運ばれます。GIsは、コオロギでは8対16個が同定されており、小川教授らはそれぞれのGIsが流速や方向など異なる情報を運んでいること、さらに上位神経節内で異なる軸索投射パターンを示すことを明らかにしました(図1左)。しかし、GIsによって運ばれた気流情報が脳内のどのような神経回路によって処理され、逃避行動の運動指令に変換されているのかはわかっていません。そこでこの課題に取り組むため、千田さんはコオロギの脳から細胞内記録を実施し、気流応答特性を解析するとともに、一つ一つの気流応答性ニューロンの形態を詳細に調べました(図2)。
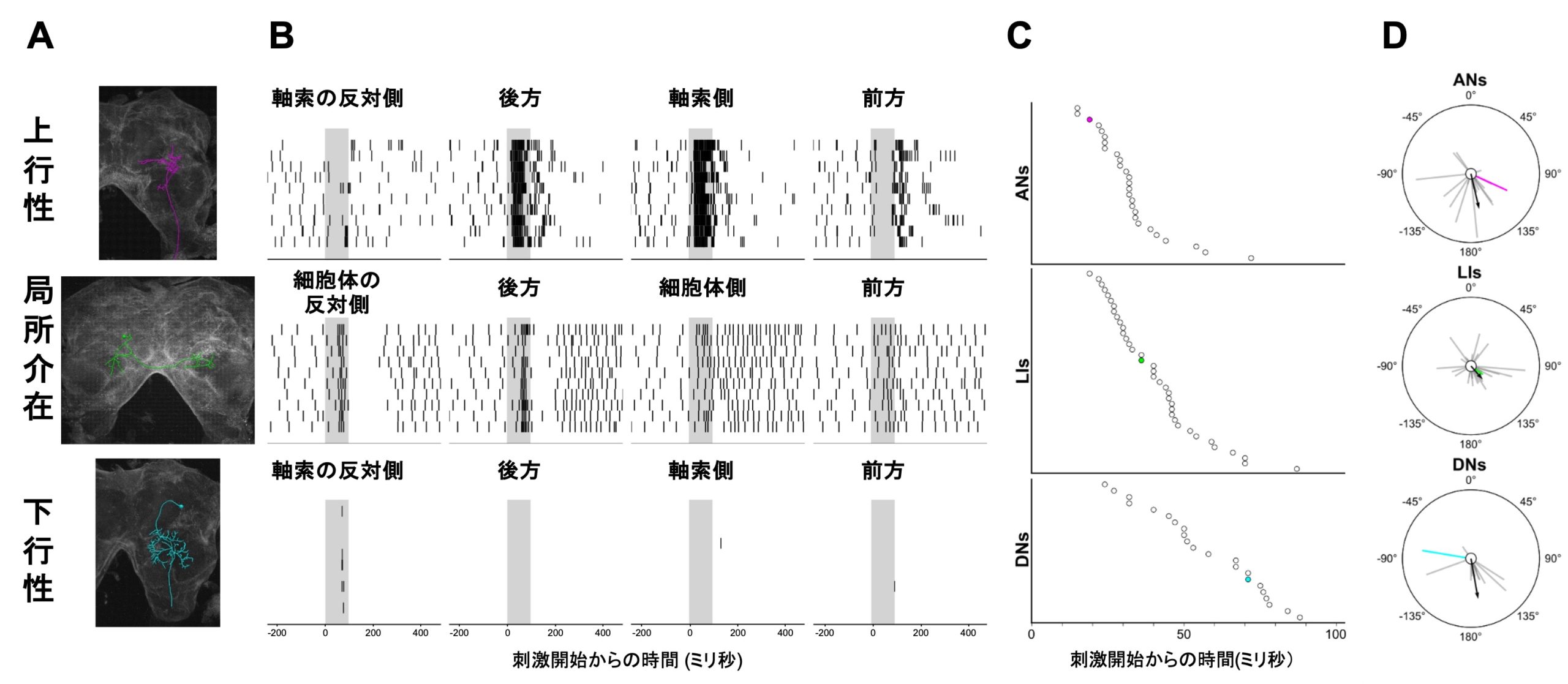
図2 コオロギ脳内の気流応答性ニューロンと反応特性 A:半側性上行性ニューロン(上)、両側性局所介在ニューロン(中)、半側性下行性ニューロンの一例。B:Aの細胞から記録された4方向からの気流刺激に対する反応を示すスパイクラスタープロット。各行は1試行の反応、縦バーはスパイク発火のタイミングを示す。グレーの影は気流刺激時間を表す。C:記録した全ての細胞における最短の応答潜時。潜時が短いものから順に並べており、色の付いたドットはAの細胞のデータを示す。D:全ての細胞における方向感受性。中心から伸びるグレーもしくは色つきの線は、4方向の刺激に対する反応強度を示すベクトルから算出した集合ベクトルで、線の方位は選好方向を、線の長さは選択性の鋭さを示す。黒矢印はそれぞれの細胞タイプのすべての選好方向ベクトルの集合ベクトルを示す。0ºは前方、180ºは後方、90ºと–90ºは上下行性ニューロンでは軸索の、局所介在ニューロンでは細胞体の同側と反対側からの刺激を示す。
今回の論文では特にGIsの軸索が投射している前大脳の腹側領域を目標として網羅的な記録を行い、最終的に92個の気流応答性ニューロンの同定に成功しました。これらのニューロンは、その形態から上行性ニューロン(腹側縦連合神経束から軸索が上行して脳内に軸索終末と軸索分枝のみが観察され、細胞体が脳にない細胞)、局所介在ニューロン(脳内に細胞体が存在し、その構造がすべて脳内に収まっている細胞)、下行性ニューロン(脳内に細胞体が存在し、その軸索を腹側縦連合神経束へ伸ばしている細胞)に分類され、さらに半側性(構造が脳の片側半球内に収まっている)細胞と両側性(構造が正中線を越えて脳の両側半球に及んでいる)細胞にそれぞれ分けられました。27個の上行性ニューロンは、一つを除いて、すべて上行性軸索と同側半球内に軸索が終末する半側性の形態を示しました。一方、42個の局所介在ニューロンのうち19個は半側性、23個は両側性で、下行性ニューロンは23個のうち15個が半側性、8個が両側性の細胞でした。
また、コオロギの前後左右の4方向から気流刺激を与え、それぞれの反応潜時と方向選択性を解析したところ、上行性ニューロンは、一つを除いて全て短い潜時の興奮性の反応を示し、特に上行する軸索と同じ側からの側方刺激に対して最も大きな反応を示しました。一方、局所介在ニューロンは35個が興奮性、7個が抑制性の応答を示し、その反応潜時や方向選択性は多様で、その形態的特徴と明瞭な相関はみられませんでした。下行性ニューロンは17個が興奮性、6個が抑制性の反応を示し、それらの反応潜時はより長く、方向選好性は様々でした。しかし、そのうち側副葉(Lateral accessory lobe)に樹状突起を広げるいくつかの下行性ニューロンは強い方向選択性を示しました。
以上の結果は、気流機械感覚情報が階層的に処理され、特に側副葉で指向性運動指令を出力する脳内神経回路構造が存在することがわかりました。現在、小川教授らは行動中のコオロギからの細胞内記録実験を進めており、これらのニューロンの活動が気流誘導性逃避行動にどのように関与するのかを明らかにしていきたいと考えています。
発表論文
Hikaru Chida, Hisashi Shidara, Hiroto Ogawa (2025)
Neural circuit architecture and directional information processing of airflow stimuli in the cricket brain. J. Neurophysiol., 134, 1214-1231.
https://doi.org/10.1152/jn.00254.2025